Estas son notas rápidas y esquemáticas para recordar algunas cosas que olvidé del neurotransmisor GABA.
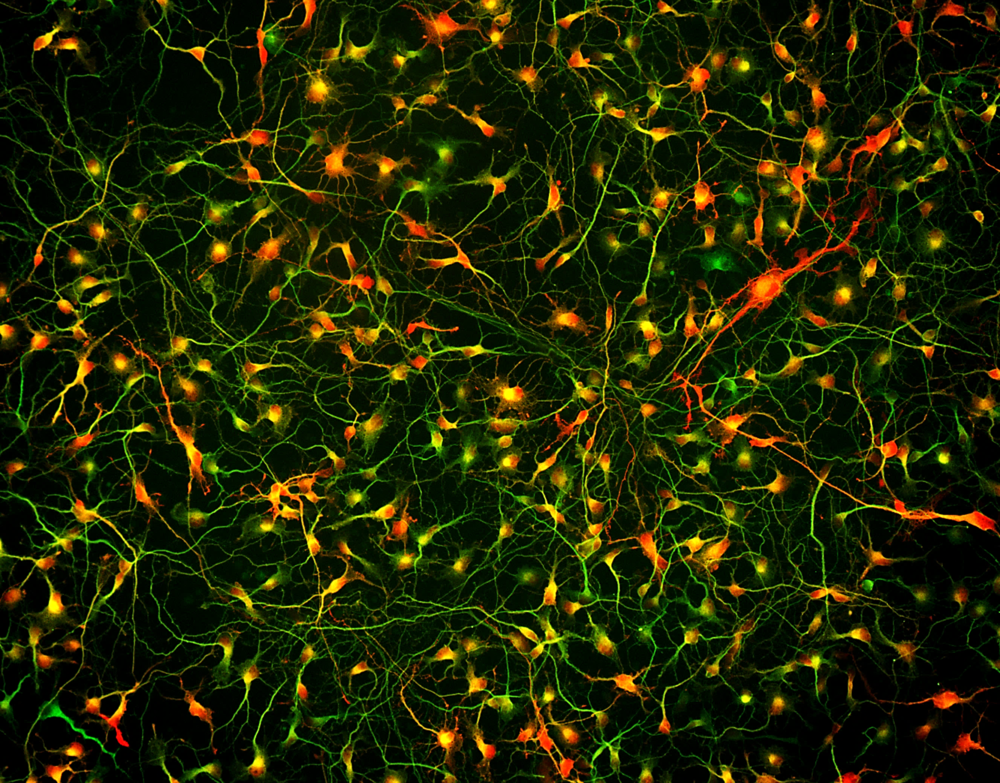
El ácido γ-aminobutírico (GABA) es el principal neurotransmisor inhibitorio en el cerebro humano. Dependiendo de la región cerebral, entre un 20% y un 40% de todas las neuronas son neuronas GABAérgicas. Los dos tipos principales de receptores GABA son el receptor ionotrópico GABA$_A$ y el receptor metabotrópico GABA$_B$.
Los receptores ionotrópicos son canales iónicos que permiten un flujo rápido de corriente y, en consecuencia, una rápida hiperpolarización o despolarización. Los receptores metabotrópicos (o acoplados a proteínas G), en cambio, no son canales iónicos sino desencadenantes de una cascada de mensajes. En particular, cuando un neurotransmisor se une a ellos, disparan una serie de señales intracelulares que involucran a la proteína G y a múltiples segundos mensajeros, lo cual eventualmente conduce a la apertura o cierre indirecto de ciertos canales iónicos. Esta cascada de señalización puede tardar cientos de milisegundos o minutos y, por lo tanto, es lenta.
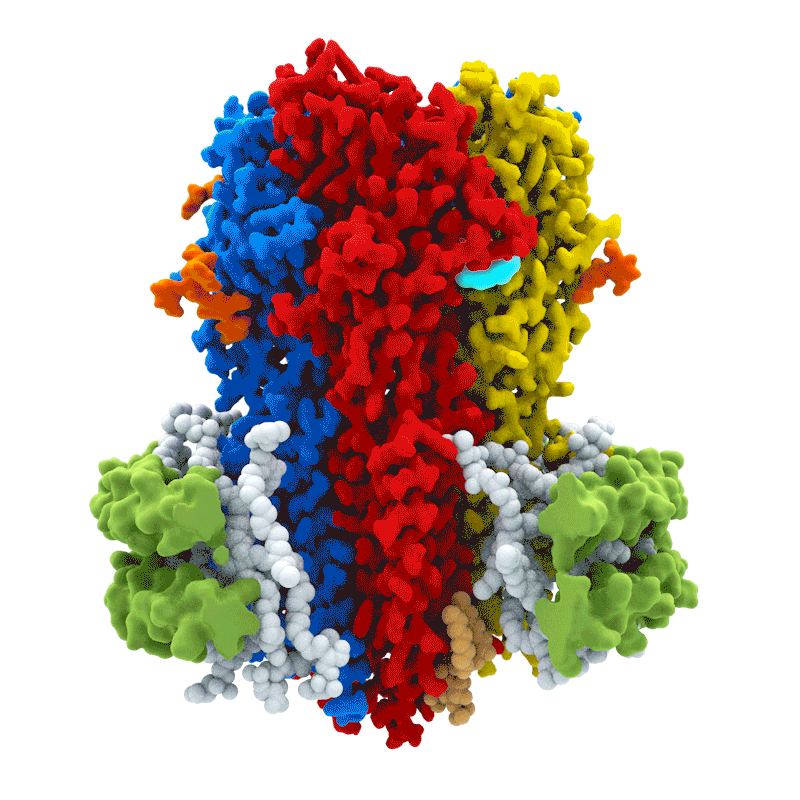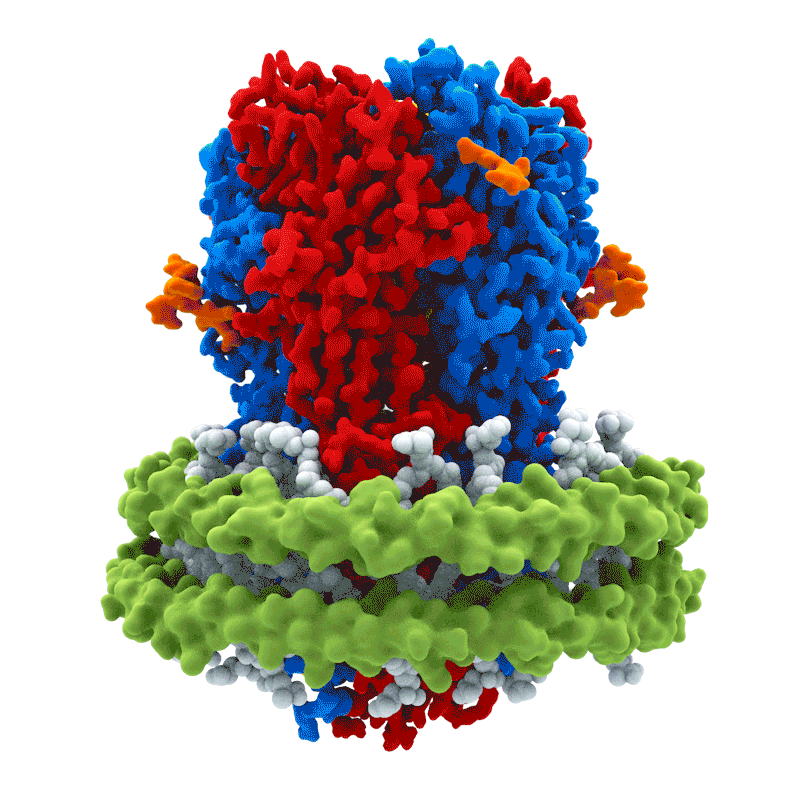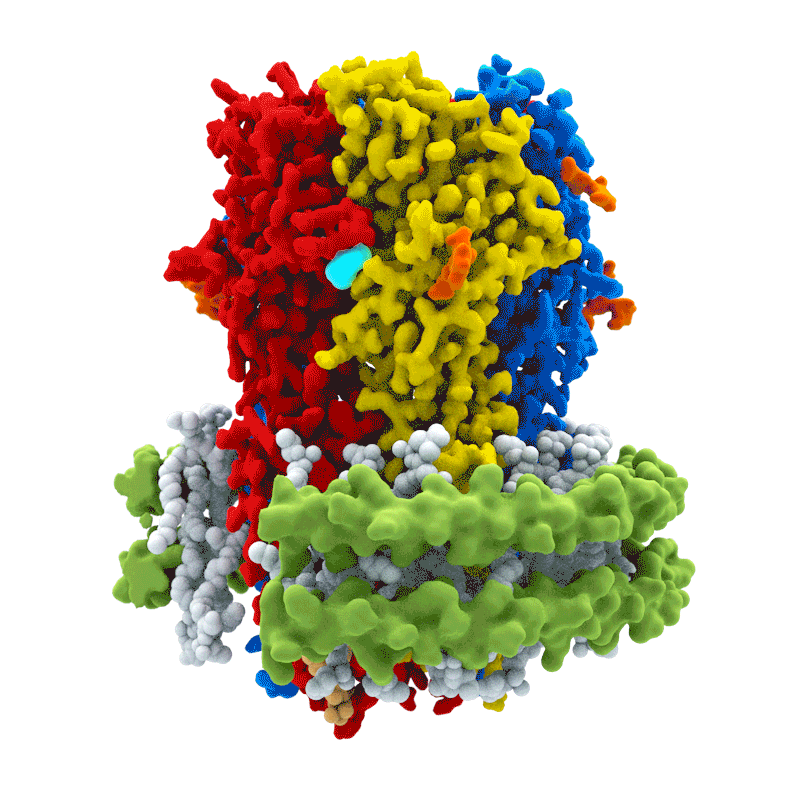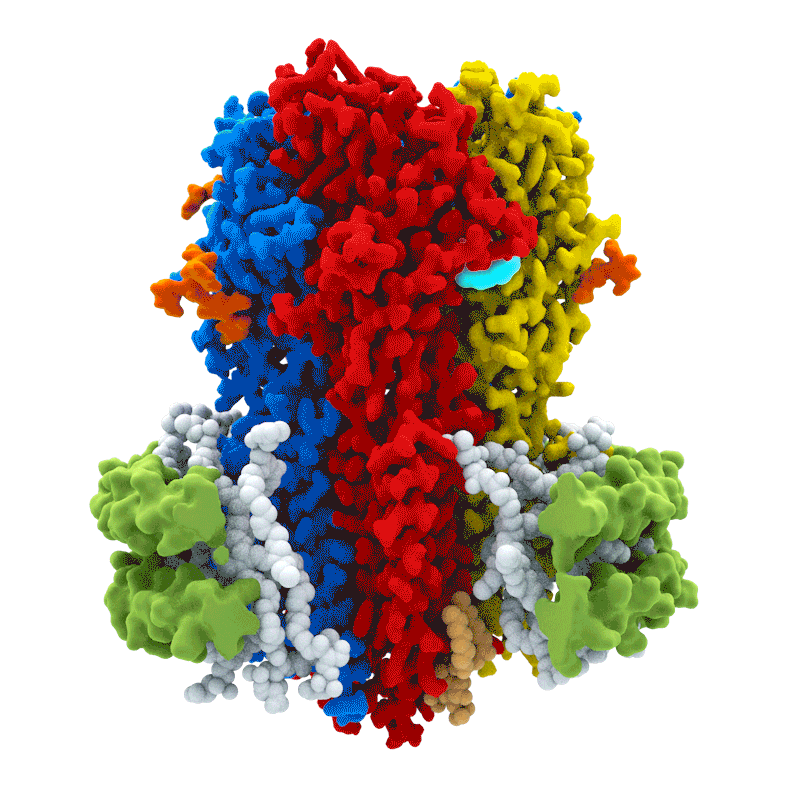
La activación de los receptores GABA$_A$ es el principal mecanismo de acción del neurotransmisor GABA. La unión al receptor GABA$_A$ permite la difusión de Cl$^-$ hacia el interior de la célula, lo que produce una hiperpolarización de la neurona postsináptica. El receptor GABA$_A$ es heterogéneo: está compuesto por subunidades $\alpha, \beta, \gamma$ y $\delta$, y existe diversidad dentro de cada familia de subunidades.
El receptor GABA$_B$ está compuesto por dos subunidades y se encuentra acoplado a canales de $K^+$ o Ca$^{2+}$ a través de una proteína G. La cascada de señales iniciada por la proteína G conduce a la activación de los canales postsinápticos de K$^+$ o a la inhibición de los canales presinápticos de Ca$^{2+}$, donde la disminución en la conductancia de Ca$^{2+}$ reduce la liberación de neurotransmisores por parte de la neurona presináptica.
Nota de color. Los receptores ionotrópicos GABA$_A$ son la clave para explicar el efecto ansiolítico de las benzodiacepinas y los barbitúricos. Cada uno de estos fármacos se une a sitios correspondientes en la cara externa del canal GABA$_A$. Si bien producen pocos cambios directos sobre el canal en sí, en presencia de GABA las benzodiacepinas aumentan la frecuencia de apertura del canal, mientras que los barbitúricos aumentan su duración. Ambos efectos resultan en un incremento del flujo de Cl$^-$ hacia el interior de la célula.
Alteraciones en los niveles del neurotransmisor GABA parecen estar relacionadas con trastornos como la esquizofrenia, la depresión y la ansiedad. Por ejemplo, la esquizofrenia se caracteriza por una densidad mayor de lo normal de neuronas en la sustancia blanca superficial/intersticial, muchas de las cuales son GABAérgicas, y una densidad reducida de estas mismas neuronas en la sustancia gris cortical (ref). En la depresión, una hipótesis etiológica digna de consideración es la así llamada GABAergic deficit hypothesis. Más allá de qué tan recisivo sean las alteraciones en el sistema GABAérgico en términos etiológicos, es algo casi establecido que existe una reducción en la actividad GABAérgica característica de la enfermedad. Estos déficits GABAérgicos pueden observarse en plasma, líquido cefalorraquídeo y tejido cortical resecado de individuos con depresión (ref), y es sugerido también por disminuciones en los niveles de inhibición intracortical en estudios con estimulación magnética transcranial.